

文献解读共生菌-宿主免疫串扰塑造硬骨鱼粘膜稳态4
人们普遍认为,当病原体克服宿主的物理/化学屏障和先天免疫时,适应性免疫是宿主的另一道防线。 通过与复杂且动态的微生物群体(共生微生物和致病微生物)的不断接触,粘膜适应性免疫已经进化出机制,可以根据细菌的有益或致病特性来识别和响应细菌的耐受性或反应性。 在过去的几年里,许多研究集中在细菌在调节适应性免疫系统中的作用以及抗原特异性适应性免疫反应对共生细菌和哺乳动物宿主之间的互惠关系的影响。 然而,尽管硬骨鱼是最古老的脊椎动物,具有由免疫球蛋白 (Igs)、B 细胞和 T 细胞组成的适应性免疫系统,但人们对这些生物体对微生物的免疫反应知之甚少。
1. 分泌型 Ig 和 B 细胞对共生细菌的反应
B细胞的主要功能是产生和分泌Ig,Ig在体液适应性免疫反应中发挥关键作用。 由于粘膜表面的特殊性质,脊椎动物在粘膜中进化出了专门的 Ig。 在哺乳动物中,分泌型 IgA (sIgA) 是粘膜表面产生的主要抗体同种型,它优先结合与宿主粘膜免疫密切相关的共生病原菌。 这一现象表明,sIgA 还可能在粘膜表面的免疫排斥、保护宿主免受侵入性病原体侵害以及调节哺乳动物宿主的共生菌群组成方面发挥重要作用。 IgT 是哺乳动物 IgA 的功能类似物,已被证明在硬骨鱼中具有独特的粘膜功能。 分泌型 IgT (sIgT) 在覆盖肠道、皮肤、鳃、口腔粘膜和咽粘膜的骨粘膜表面含量丰富。 共生细菌的主要 Ig 同种型,表明 sIgT 在硬骨鱼粘膜微生物群稳态中起主导作用。 最近,新开发的 IgT-seq 技术证明了 sIgT 覆盖鳃中有益细菌的能力,包括已知产生短链脂肪酸 (SCFA) 的几种分类群,例如具有抗炎特性的乳杆菌和梭杆菌。图1)。

图 1. 硬骨鱼中微生物群落及其化合物对宿主稳态和对微生物群落的粘膜适应性免疫的影响概述。
说明:在三种硬骨鱼 Ig 中,sIgT 是粘液中覆盖微生物的主要 Ig,而 IgT 靶向微生物包括产生具有抗炎特性的短链脂肪酸 (SCFA) 的类群。 sIgM靶点与sIgT的共生性较低,对致病菌和非致病菌的生长都有抑制作用。 相比之下,sIgD 被证明与微生物驱动的克隆扩张和多样性一致,这可能会影响共生细菌的粘附。 抗体分泌细胞 (ASC) 在细菌反应中的作用已得到证实,但硬骨鱼粘膜中 B 细胞的吞噬能力仍不清楚。 当一些机会性病原体入侵时,IgT+B细胞迅速增殖并产生特异性IgT。 同时,T细胞产生细胞因子,如TNFα和IFNγ,参与对不同细菌刺激的反应。 众所周知,产生鞘脂的细菌类群可以诱导持续的 IgT 表达和分泌; 关键是益生菌促进T细胞增殖,提高Ig水平。 此外,Rag1基因敲除的斑马鱼比幼鱼表现出更高的弧菌种类丰度,而T细胞的持续转移降低了弧菌种类的生长和多样性。 此外,没有 IL-17/F1 的鱼表现出明显更高的致病细菌丰度。
然而,IgT 针对微生物(包括共生菌和病原菌)的健康促进或诱发疾病作用还需要进一步验证。 在哺乳动物中使用 IgA-seq,从炎症性肠病 (IBD) 或脊柱炎患者中分离出的 IgA 高肠道细菌分类群被确定与疾病发病机制相关。 有趣的是,将这些精选的纯化 IgA 靶向细菌菌株转移到无菌小鼠模型中会导致对肠病、结肠炎或炎性关节炎的显着易感性,而从健康个体中分离出的高 IgA 涂层细菌菌株则表现出保护小鼠的能力对抗炎症性疾病。 此外,在 IgT 耗尽后发现虹鳟鱼鳃相关细菌类群的丰度增加。 由于缺乏sIgT,大量共生细菌转移到鳃片,血液中发现大量脂多糖(LPS)。
总而言之,sIgT 对于 sIgT 微生物群落在鳃粘膜层内的定殖、锚定和遏制是必需的(图 2)。 同样,缺乏 IgA 或粘膜聚集 Ig 受体的小鼠表现出肠道共生菌失调的多种迹象,包括细菌丰度减少、微生物组成失衡、对肠道炎症疾病的高度易感性以及明显的微生物敏感性。 少量。 有趣的是,sIgA 可以通过多种机制覆盖共生细菌,包括典型的 Fab 依赖性抗原特异性细菌识别、非典型的 Fab 独立性和非抗原特异性聚糖介导的相互作用,以及通过微生物群编码的超抗原靶向 sIgA。 然而,硬骨鱼中 sIgT 与微生物结合的机制和功能意义仍然知之甚少。
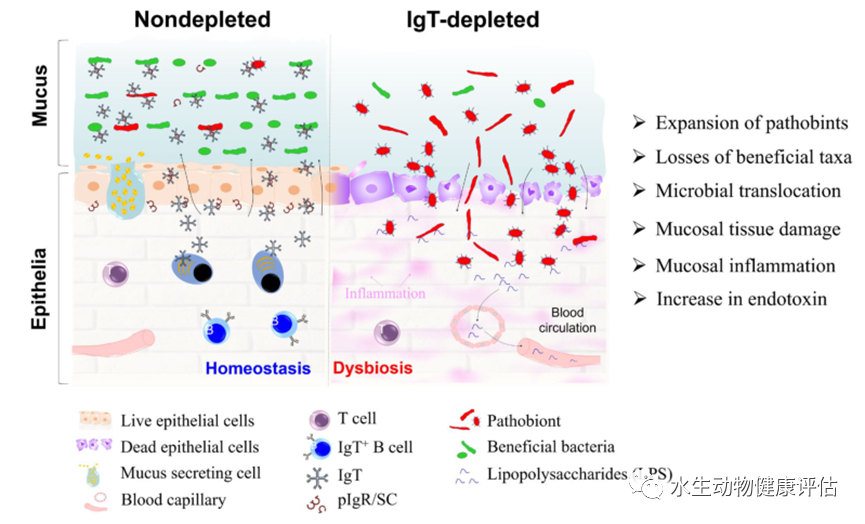
图2 IgT在硬骨鱼粘膜稳态中的作用
说明:正常情况下,鱼类体内IgT+B细胞产生的sIgT通过pIgR通过上皮细胞转运至粘液层,pIgT可以覆盖大量含有病原菌和有益菌的微生物群落,实现体内平衡。 然而,这种平衡在 IgT 缺陷的鱼类中被破坏,微生物群发生严重的菌群失调,其特征是病原微生物的扩张和有益类群的丧失。 同时,一些机会致病菌从粘液层转移到上皮,引起明显的粘膜组织损伤和炎症。 此外,IgT 缺陷鱼血清中的 LPS 水平显着升高,表明细菌已进入体循环。
此外,硬骨鱼中的IgM是血浆中的主要Ig,已被确定为全身免疫的主要介质。 粘液中高 IgM 蛋白浓度表明这种 Ig 也可能参与粘膜免疫反应。 先前的研究表明,虹鳟鱼 IgM 可以用共生细菌覆盖粘膜表面。 尽管如此,虹鳟鱼 IgM 在维持微生物群稳态中的功能仍然未知。 最近的研究表明,河豚IgM重链分子作为N-乙酰氨基葡萄糖结合蛋白,对革兰氏阳性菌和革兰氏阴性菌,包括致病菌和非致病菌的生长具有显着的抑制作用,表明IgM具有消除虹鳟鱼中细菌的能力。 IgD 是一种古老的抗体,在硬骨鱼的所有免疫组织中表达,然而,分泌型 IgD (sIgD) 的生物学作用仍然是一个有争议的话题。 有趣的是,sIgD 可以与肠道共生菌群结合,进而影响 IgD 基因的转录水平。 sIgD 可以影响共生细菌对肠上皮的粘附并限制病原体的侵入。 在哺乳动物中,这一过程通常与 IgM 和 IgA 之间的合作有关。 因此,可以推测sIgD活性的机制可能涉及IgD和IgM的协同作用。
2. T细胞对共生细菌的反应
T细胞与B细胞合作,在细胞介导的适应性免疫中发挥关键作用,通常还与粘膜细菌相互作用,诱导对共生菌群各种成分的耐受或免疫,以维持粘膜稳态。 状态。 在脊椎动物中,表达 αβ T 细胞受体 (TCR) 的 T 细胞是最大的 T 细胞亚群,除了通常分别与哺乳动物相关的两个主要共同受体 CD4 和 CD8 之外。 T 辅助细胞 (Th) 和 T 细胞毒性 (Tc) 细胞相关。 在硬骨鱼中,一些物种的肠道中发现了类 T 细胞群,例如大西洋鲑鱼、鲈鱼、鲤鱼和虹鳟鱼。 在斑马鱼中,肠道 T 细胞最近被证明可以通过调节弧菌物种多样性来调节共生细菌的组成。 布鲁格曼等人。 报道称,肠道微生物群中的弧菌种类比斑马鱼发育早期(此时适应性免疫尚未存在)以及缺乏适应性免疫反应的 Rag1 敲除斑马鱼更丰富。 幼稚的成年斑马鱼,表明先天免疫系统无法控制肠道中的所有共生成员。 然而,将 T 淋巴细胞过继转移到 Rag1 敲除斑马鱼中会降低弧菌物种的生长和多样性,这表明 T 细胞在维持肠道微生物稳态中发挥着关键作用。 然而,体内 T 细胞介导的抑制机制尚不清楚,Brugman 等人。 研究发现,在体外暴露于 Rag1 敲除鱼共生体的 T 细胞能够产生并分泌更多的 IFNγ,这表明 T 淋巴细胞可能通过诱导炎症反应在调节粘膜细菌中发挥重要作用。 因此,未来的研究可以集中于阐明T细胞是否被细菌配体激活,以及阐明记忆T细胞在弧菌物种免疫激活和抑制中的作用。
3. 共生菌群形成粘膜适应性免疫反应
先前的研究已经阐明了细菌细胞壁成分和共生微生物代谢物的免疫调节作用,这些成分影响哺乳动物肠粘膜中的各种细胞类型,包括 B 和 T 淋巴细胞。 例如,无菌小鼠肠道相关淋巴组织 (GALT) 缺乏成熟,适应性 T 或 B 细胞很少,并且适应性体液反应存在广泛缺陷。 然而,这些免疫相关的细胞和分子成分在共生微生物定植后迅速积累,并立即渗透到肠固有层效应位点。
然而,尽管适应性免疫系统的所有组成部分,包括 T 细胞和 B 细胞,都存在于斑马鱼发育的早期阶段,但它们直到受精后 4-6 周才达到免疫能力并诱导有效的适应性免疫。 反应。 因此,斑马鱼在发育的前 30 天左右完全依靠其先天免疫反应来避免病原体感染,并且尚未开发出将无菌斑马鱼培养至成年的适当方法。
由于硬化粘膜表面不断与水生环境接触,因此它们含有大量机会性细菌病原体,如果宿主受到缺氧、温度变化、高密度、和机械损伤。 可能会导致疾病。 因此,更好地了解机会病原体与宿主免疫系统之间的相互作用至关重要。 未来的研究应重点关注针对机会性细菌病原体的特定 Ig 防御机制。
参考:
于YY,丁LG,黄ZY,等。 共生细菌-免疫串扰影响硬骨鱼粘膜稳态[J]. 水产养殖评论,2021,13(4):2322-2343。
土井:doi.org/10.1111/raq.12570

结尾
公众号邮箱:ehbiotech@163.com
(竭诚欢迎您的投稿、建议或合作联系)

如果你觉得这样更好看,请点击这里↓↓↓






